Quaderns de Psicologia | 2022, Vol. 24, Nro. 2, e1731 | ISNN: 0211-3481 | ![]()
 https://doi.org/10.5565/rev/qpsicologia.1731
https://doi.org/10.5565/rev/qpsicologia.1731

Orden espacial de los eventos y distribución de la actividad en la función suplementaria
Spatial order of events and distribution of activity in the supplementary function
Felipe de Jesús Patrón Espinosa
Universidad Autónoma de Baja California
Carlos de Jesús Torres Ceja
Carlos Javier Flores Aguirre
Universidad de Guadalajara
Mauricio Ortega González
Universidad Autónoma de Baja California
Resumen
Se exploran los efectos de variar el orden espacial de la disponibilidad de producción del agua sobre las trayectorias empleadas por el organismo y la ubicación espacio-temporal de sus respuestas (introducción de cabeza al bebedero y de presión de palanca). Dos grupos de cuatro ratas fueron expuestos en cámaras de condicionamiento ampliadas a condiciones en las que la disponibilidad de producción varió en tres palancas. Para el Grupo 1, en la Fase 1 la disponibilidad se limitó a una palanca, en la Fase 2 varió en orden secuencial en tres palancas. En la Fase 3, la disponibilidad varió aleatoriamente en tres palancas. El Grupo 2 fue expuesto a fases similares, pero en un orden inverso. Los resultados mostraron que las ratas reprodujeron los parámetros espaciales programados para la producción del agua. Se observó un efecto de arrastre consistente entre ratas, lo que se interpretó como desligamiento funcional.
Palabras clave: Teoría de la conducta; Función suplementaria; Conducta espacial; Ratas
Abstract
We explore the effects of varying the spatial order of water production availability on the trajectories employed by the organism and the spatio-temporal location of its responses (head introduction into the liquid dispenser and lever pressure). Two groups of four rats were exposed in expanded conditioning chambers to conditions in which production availability varied by three levers. For Group 1, in Phase 1 availability was limited to one lever, in Phase 2 it varied in sequential order across three levers. In Phase 3, availability varied randomly across three levers. Group 2 was exposed to similar phases but in a reverse order. The results showed that the rats reproduced the spatial parameters programmed for water production. A consistent carryover effect was observed between rats, which was interpreted as functional detachment.
Keywords: Behavior theory; Supplementary function; Spatial behavior; Rats
Introducción
Diversos autores han señalado que, a lo largo de la historia de la psicología experimental, la investigación con animales no humanos se ha caracterizado por brindar mayor atención a la dimensión temporal de los fenómenos psicológicos por encima de la dimensión espacial (Christie, 1996; Leising et al., 2014; McClearn y Harlow, 1954; Nasser y Delamater, 2016; Pavlov, 1928; Rescorla y Cunningham, 1979; Ribes, 1992). A pesar de esto, al realizar una revisión minuciosa de fuentes académicas es posible encontrar investigaciones que se han interesado por la dimensión espacial del comportamiento animal, en algunas ocasiones concentrándose en su estudio y en otras incluyendo a la dimensión temporal. Focalizando la atención en la dimensión espacial, estos estudios podrían categorizarse de la siguiente forma:
(1)Experimentos con el objetivo de estudiar las propiedades espaciales de la actividad del organismo, limitando la dimensión espacial a los cambios en la actividad como efecto de manipulaciones experimentales.
(2)Experimentos con el propósito de conocer los efectos de manipular cierta propiedad espacial de los eventos a los que es expuesto el organismo, limitando la dimensión espacial los cambios programados por el investigador en la situación experimental.
(3)Experimentos en los que se combinan los dos objetivos previos, es decir, se busca conocer los efectos de manipular propiedades espaciales de la preparación experimental sobre propiedades espaciales de la actividad del organismo.
En lo que respecta a la Categoría 1, una investigación clásica que podría incluirse fue realizada por Tamara Dembo (1930) con el propósito de estudiar en una situación “abierta” la conducta dirigida hacia metas en ratas. Para esto, se empleó una cámara experimental de 70 por 120 cm que se encontraba segmentada por dos paneles o paredes de vidrio que posibilitaban la visualización de la meta (alimento) en una ubicación espacial fija. El análisis de datos se basó en las trayectorias, revelando que, para las ratas con nula experiencia, éstas fueron erráticas, pero con pocos ensayos las trayectorias mostraron direccionalidad hacia la meta. Estudios más recientes que pueden agregarse en la Categoría 1 se caracterizan por explorar los efectos de diversos programas de reforzamiento sobre la distribución espacial de la actividad de los organismos. Cabe señalar que en estos estudios, aunque pueden estar activos simultáneamente más de un programa en distintos dispositivos, así como variar la probabilidad de su activación, su ubicación espacial no varía (Baum y Rachlin, 1969; Pear, 1985; Pear y Rector, 1979).
En la Categoría 2 es posible incluir el estudio clásico realizado por Edward Tolman et al. (1946) para responder si las ratas, al desplazarse en situaciones experimentales como los laberintos T, toman ciertas direcciones debido a que, a través del entrenamiento, desarrollan una disposición a responder ante un estímulo en particular o si, de forma más compleja, desarrollan una disposición a orientarse hacia un lugar. Para poner esto a prueba, se empleó un laberinto en forma de equis que contaba con dos posiciones de salida contrapuestas en los extremos de un pasillo y dos metas contrapuestas en los extremos del otro pasillo. Un grupo de ratas fue entrenado para encontrar alimento siempre que presentara la respuesta de girar hacia la derecha, mientras que el segundo grupo fue entrenado para ir siempre al mismo lugar-meta independientemente del lugar de salida. Los resultados en la fase de prueba mostraron que las ratas del segundo grupo dejaron de presentar errores al elegir el brazo-meta antes del Ensayo 8, mientras que la mayoría de ratas del primer grupo continuó presentando errores hasta el Ensayo 72. Ambos hallazgos llevaron a los autores a concluir que las ratas desarrollaron una disposición a orientarse a un lugar más que una disposición de presentar una respuesta ante un estímulo particular.
Dentro de la Categoría 2, se podrían agregar estudios más actuales en los que, desde el paradigma operante, se trata de identificar los efectos de variar la contigüidad espacial entre el lugar en el que se presenta la respuesta procuradora y la entrega del reforzador sobre la tasa de respuesta (Leising al., 2014). En esta categoría también pueden incluirse los estudios que se interesan en los efectos de variar el tamaño de la cámara experimental sobre la tasa de respuesta (Skuban y Richardson, 1975).
En relación con la Categoría 3, un estudio clásico fue desarrollado en 1946 por Petr Kupalov con el objetivo de poner a prueba los hallazgos que se tenían hasta el momento sobre el condicionamiento pavloviano, concentrándose en la actividad motora condicional presentada por perros en situaciones “libres”. Para esto, se empleó un cubículo de 7.5 x 5 m que contó con diversas ubicaciones en las que podían presentarse estímulos neutros como metrónomos y estímulos incondicionales (EI), así como una alfombra con la que se marcaban zonas específicas. La presentación de los estímulos podía variar de ubicación y el inicio del ensayo dependía de que el perro se ubicara en una zona en particular. Se encontró que la zona asociada a la presentación de los estímulos adquiere propiedades apetitivas cuando el EI es apetitivo. Por su parte, si otra zona del cuarto es asociada con la aplicación de un estímulo condicionado inhibitorio o con la extinción del condicionamiento excitatorio, la zona adquiere propiedades inhibitorias. Kupalov concluyó que la zona en conjunto con el estímulo condicional se convierte en un factor que controla la conducta voluntaria del perro (como se citó en Kupalov et al., 1964).
Entre los estudios más recientes que podrían incluirse en la Categoría 3 se encuentran los que han empleado el laberinto acuático de Morris para conocer los efectos de variar la distancia espacial entre las señales y la meta sobre el aprendizaje espacial (Chamizo y Rodrigo, 2004; Maes et al., 2009; Rodrigo et al., 2014). También podrían agregarse los estudios que emplean la tarea de aprendizaje tiempo-lugar (ATL) que, en términos generales, implica la exposición de los organismos a una situación en la que se presentan estímulos motivacionalmente significativos en distintas ubicaciones, por ejemplo, en las cuatro paredes de la cámara experimental. La ocurrencia en tiempo y espacio de estos estímulos puede ser aleatoria o en un orden consecutivo (Carr y Wilkie, 1998; Reebs, 1996; Saksida y Wilkie, 1994).
Concentrando la atención en la psicología interconductista (Kantor, 1959), diversos autores han llamado la atención acerca de incluir el análisis de la dimensión espacial en los estudios empíricos (Irigoyen et al., 1999; Ray y Brown, 1975; Ribes, 1992). Específicamente, desde Teoría de la conducta (Ribes y López, 1985; Ribes, 2010), el estudio de esta dimensión ha tomado, al menos, dos posibles direcciones: la primera, incluida en la Categoría 1, al centrarse en el desarrollo de medidas o modos de representación (Camacho, 2017) y la segunda que se incluye en la Categoría 2 al centrarse en conocer los efectos de manipular propiedades espaciales de los eventos sobre el comportamiento. Respecto de la primera dirección, los estudios se han centrado en explorar los efectos de diferentes programas de reforzamiento presentados de forma concurrente en cajas de condicionamiento ampliadas (Ribes et al., 2020; Ribes y Tamayo, 2020). Al parecer, el propósito principal de estos estudios ha sido analizar el comportamiento animal empleando medias molares (Ribes, 2007) desde posicionamientos teóricos nuevos (Ribes, 2018; Ribes, 2020).
Por su parte, en la segunda dirección podrían incluirse estudios como el realizado por Felipe Patrón et al. (2014), en el que exploraron los efectos de variar la distancia espacial entre los elementos que integran la función suplementaria (Ribes y López, 1985) sobre el tiempo entre respuestas de presión de palanca y el porcentaje de aguas producidas. Para esto, se emplearon dos cámaras de condicionamiento ampliadas en las que fue posible instalar distintos dispositivos de estimulación en diferentes zonas de la pared operativa. Para un primer experimento, las ratas fueron expuestas a condiciones en las que varió la distancia entre la palanca de respuesta y el conjunto luz-bebedero. En un segundo experimento, se varió la distancia entre el bebedero y el conjunto luz-palanca. Los autores encontraron que aumentar la distancia espacial entre los dispositivos de estimulación generó mayor variabilidad en el tiempo entre respuestas, así como porcentajes de aguas producidas más bajos; esto de forma más pronunciada al variar la ubicación del bebedero.
El estudio elaborado por Patrón et al. (2014) podría incluirse en la Categoría 2 para estudios sobre la dimensión espacial, pues manipularon propiedades espaciales de los eventos que constituyen la situación experimental, pero no describieron las propiedades espaciales de la actividad de los organismos. Continuando con esta línea de investigación, se pretende que el presente estudio cumpla con los requisitos para ser incluido en la Categoría 3, al contar con el objetivo de explorar el efecto de variar el orden espacial de la disponibilidad de producción del estímulo contextualizador (agua) sobre las trayectorias empleadas por el organismo y la ubicación espacio-temporal de sus respuestas (introducción de cabeza al bebedero y de presión de palanca).
Método
Sujetos
Se emplearon ocho ratas hembras de la cepa Wistar, de tres meses de edad, ingenuas y con un peso corporal, de entre 189 y 275 g al inicio del experimento. Todas éstas se mantuvieron bajo un régimen de privación de agua de 23 horas y 25 minutos al día. Después de cada sesión experimental tuvieron acceso libre y de forma individual a bebederos con agua durante cinco minutos en sus cajas habitación que se ubicaban en una colonia con temperatura (23-25 grados centígrados) y humedad controlada (45 %-55 %), así como ciclos día/noche de 12 horas. El alimento se encontró disponible mientras que las ratas se encontraron en sus cajas habitación. Las sesiones experimentales se llevaron a cabo todos los días de la semana exceptuando domingo.
Aparatos
Se empleó una cámara experimental de 85 cm de largo por 42 cm de ancho y 36 cm de alto con la opción para la instalación de distintos módulos de estímulo y respuesta en tres puertos ubicados en su pared operativa. El puerto central se encontraba a la mitad de la pared larga (85 cm) opuesta a la puerta por la que se introducían los sujetos. Los otros dos puertos (sur y norte) se encontraban a los lados del puerto central a una distancia de 15 cm. En cada puerto se pudo instalar, dependiendo de la fase experimental: (a) un estímulo visual que constó de una tecla translúcida de 2,5 cm que podía iluminarse de color amarillo por medio de un foco de 28-V ubicado a 9 cm del suelo, (b) un dispensador de agua de tipo gotero que entregó 0,3 cm3 de agua en cada activación (marca MED modelo ENV-201A) ubicado a 2 cm del suelo y (c) una palanca de respuesta (marca MED modelo ENV-110M) ubicada a 2 cm del suelo con un criterio de 0.2 N de fuerza para cerrar su microinterruptor. Cabe señalar que, debido a que el dispensador de agua fue de tipo gotero, el agua producida no contó con un límite de tiempo para el acceso. Además, el bebedero contó con un sensor de respuestas de entrada de cabeza (marca MED modelo ENV-251L).
En los tres puertos se instalaron palancas y arriba de éstas a 9 cm del suelo se instaló un estímulo visual (tecla). Únicamente en el puerto sur se instaló a un lado de la palanca un bebedero. El registro de la ubicación espacial de las ratas se realizó con la delimitación de tres zonas (17 cm x 10 cm) del área de la cámara experimental dependiendo de su proximidad con los dispositivos de estímulo y respuesta (rectángulos 1, 2 y 3 en la Figura 1). La cámara se encontraba dentro de un cubículo de aislamiento acústico de color negro, mismo que contaba con dos ventiladores de 115W que proporcionaron ventilación y ruido blanco. La luz general fue proporcionada por un foco de luz negra-ultravioleta de 20 W. En la Figura 1 se muestran las medidas de la cámara experimental y la disposición espacial en la que se instalaron los dispositivos para la fase con mayor contigüidad espacial.
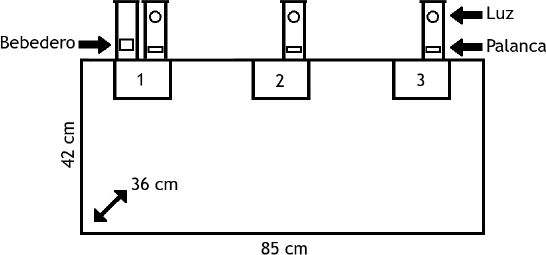
Figura 1. Descripción esquemática de la cámara experimental
Nota. Los rectángulos con los números 1, 2 y 3 representan las zonas próximas a los puertos sur, central y norte en los que pudieron instalarse los dispositivos de estimulación.
Procedimiento
Se empleó un diseño de replicación intrasujeto (Arnau, 1990) en la modalidad reversible multivariable (Castro, 1977) con tres réplicas por grupo. Todos los sujetos fueron expuestos a una fase previa al experimento que consistió en entrenamiento para producir agua al presionar una palanca. Estas sesiones finalizaron después de 60 min o si se producían 100 estímulos contextualizadores (agua). Durante este procedimiento únicamente se instaló la palanca central por debajo de un estímulo visual (tecla) y a un lado de un bebedero ubicado, también, en la zona central. Posteriormente, las ratas fueron divididas en dos grupos y expuestas a un programa que entregó agua cada 30 s sólo si el sujeto presionaba al menos una vez la palanca dentro de este ciclo de tiempo. La disponibilidad de agua fue señalada por medio de la iluminación del estímulo visual. Con la primera respuesta en la palanca que se encontraba debajo del estímulo visual iluminado, se apagaba la luz y se producía el agua al final del ciclo de 30 s.
Para la Fase 1, el Grupo 1 fue expuesto a condiciones en las que la producción de agua estuvo disponible constantemente en la primera palanca sur (adjunta al bebedero). En la Fase 2 la disponibilidad de la producción varió entre los ensayos en un orden secuencial a través de las tres palancas (palanca sur – palanca central – palanca norte). Para la Fase 3, la disponibilidad de producción se presentó aleatoriamente en alguna de las tres palancas para cada ensayo. Las fases para el Grupo 2 fueron similares a las del Grupo 1 con la diferencia de que el orden de condiciones a las que fueron expuestas las ratas fue invertido. Cada fase constó de 15 sesiones que concluían al transcurrir 60 ensayos que iniciaban cuando se colocaba a la rata en el área central de la cámara. Al finalizar la Fase 3 todas las ratas fueron expuestas a siete sesiones de reversión en las que las condiciones fueron similares a las de la Fase 1 de su respectivo grupo.
Estrategia de análisis de datos
La programación y el registro de eventos se realizó mediante un equipo de cómputo que contó con una interfase y el software MED-PC IV para ambiente Windows. El análisis y representación gráfica de los datos se realizó a través de los software TopScan 2.0® y SigmaPlot 11.0®. Una cámara Panasonic WV-CP284® fue empleada para el registro en vídeo de las sesiones experimentales.
Consideraciones éticas
El estudio se realizó cumpliendo con la NORMA Oficial Mexicana NOM-062-ZOO-1999 para la producción, cuidado y uso de los animales de laboratorio, así como con la NORMA Oficial Mexicana NOM-087-ECOL-SSA1-2002 para la clasificación y manejo de residuos biológicos.
Resultados
La Figura 2 muestra la proporción de trayectorias (ordenada) empleadas por las ratas del Grupo 1 para trasladarse entre las Zonas 1, 2 y 3 durante las primeras cinco y últimas cinco sesiones para cada fase (abscisa). Cada trayectoria implicó la entrada de la rata en las zonas de la cámara experimental para los siguientes casos, incluyendo en orden inverso: Zona 1 – Zona 2 (T 12), Zona 2 – Zona 3 (T 23), Zona 1 – Zona 2 – Zona 3 (T 123), Zona 1 – Zona 3 (T 13). En la Fase 1 se observaron mayores proporciones para el traslado de la Zona 1 a la 2 (Trayectoria 12) en todas las ratas, con excepción de las Ratas R23 y R24 que durante las primeras sesiones mostraron una mayor proporción por la Trayectoria 123 con valores que oscilaron entre 0.5 y 0.7. Para la Fase 2 no se observó alguna tendencia por emplear una trayectoria en mayor proporción, exceptuando la Trayectoria 12 para las Ratas R24 y R22 en las primeras y últimas sesiones de la fase respectivamente. En la Fase 3 las Ratas R22 y R24 mostraron una proporción mayor para la Trayectoria 12, mientras que en las demás ratas no se observó alguna tendencia por el empleo de una trayectoria. Este efecto se replicó en la fase de reversión.
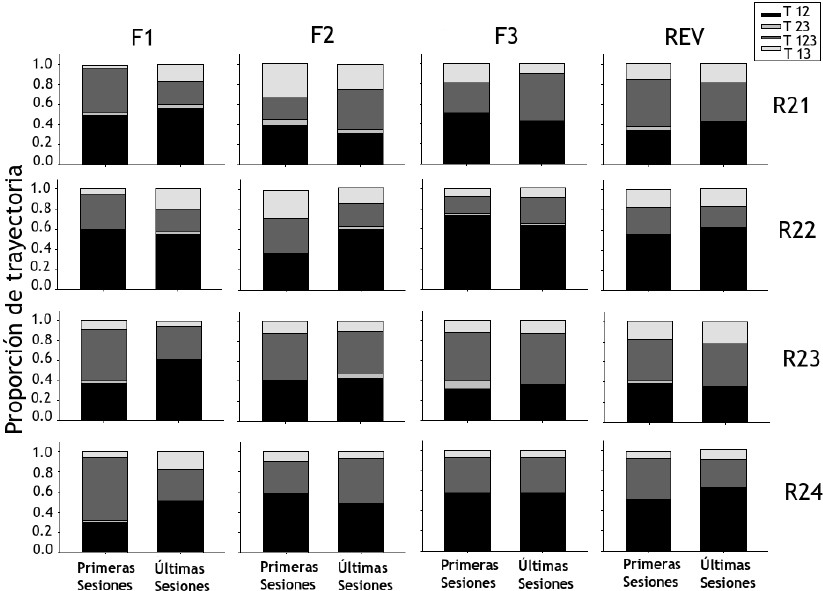
Figura 2. Proporción de trayectorias empleadas a través de las zonas próximas a los dispositivos para el Grupo 1 durante las primeras y las últimas sesiones de cada fase
Nota. Donde R es rata, T es trayectoria y F es fase.
La Figura 3 muestra la proporción de trayectorias empleadas (ordenada) por las ratas del Grupo 2 durante las primeras cinco y últimas cinco sesiones para cada fase (abscisa). Para la Fase 1, los resultados mostraron proporciones similares en las Trayectorias 12 y 123 para todas las ratas, con valores que oscilaron entre el 0.4 y el 0.5. Este efecto se replicó a través de las fases, manteniéndose las proporciones relativamente estables. La excepción fue la Rata R27 durante las Fases 3 y 4 mostrando una proporción mayor por la Trayectoria 12 con valores cercanos al 0.6.

Figura 3. Proporción de trayectorias empleadas a través de las zonas próximas a los dispositivos para el Grupo 2 durante las primeras y las últimas sesiones de cada fase
Nota. Donde R es rata, T es trayectoria y F es fase.
La Figura 4 muestra la proporción de respuestas de presión de palanca (Ryx) y de introducción de cabeza al bebedero (Rxy) para las ratas del Grupo 1 (ordenada) según los cinco subintervalos en los que se dividieron los ensayos en las primeras cinco y últimas cinco sesiones (abscisa). En relación con las Ryx, durante la Fase 1 se observó mayor proporción en los Subintervalos 3, 4 y 5 para la mayoría de las ratas. Durante la Fase 2 las proporciones se estabilizaron alrededor del valor de 0.2 para todas las ratas. En la Fase 3 este efecto se mantuvo, pero durante las primeras sesiones en las Ratas R22 y R23 se observó una proporción ligeramente mayor en los Subintervalos 4 y 3 respectivamente. Para las últimas sesiones de esta fase, la Rata R21 mostró proporciones ligeramente mayores en los Subintervalos 4 y 5, mientras que la Rata R23, en los Subintervalos 3 y 4. Durante la fase de reversión las Ratas R21 y R22 mostraron proporciones mayores de Ryx en los Subintervalos 3 y 4, mientras que las Ratas R23 y R24, en el Subintervalo 3 durante las últimas sesiones.
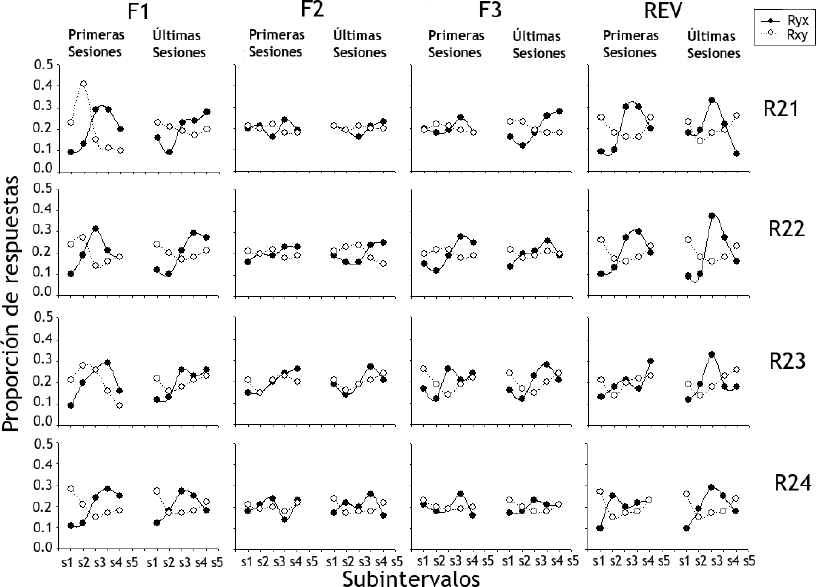
Figura 4. Proporción de respuestas de presión de palanca (Ryx) y de introducción de cabeza al bebedero (Rxy) para las ratas del Grupo 1 según los cinco subintervalos en los que se dividieron los ensayos en las primeras cinco y las últimas cinco sesiones
Nota. Donde R es igual a rata y F igual a fase.
Con respecto a las Rxy, durante las primeras sesiones de la Fase 1 se observaron proporciones mayores en los Subintervalos 1 y 2 con 0.4 como el valor más alto, pero esta diferencia no se presentó durante las últimas sesiones de la misma fase. Este efecto se mantuvo relativamente estable durante las Fases 2 y 3 con valores próximos al 0.2. En la fase de reversión las respuestas se concentraron en los Subintervalos 1 y 2 con valores cercanos a 0.25.
La Figura 5 muestra la proporción de respuestas de presión de palanca (Ryx) y de introducción de cabeza al bebedero (Rxy) para las ratas del Grupo 2 (ordenada) según los cinco subintervalos en los que se dividieron los ensayos en las primeras y últimas cinco sesiones (abscisa). Los resultados sobre las Ryx mostraron que no hubo efectos abruptos con los cambios de fases ni a lo largo del experimento. En todas las ratas, exceptuando la R25, se observaron proporciones similares para los distintos subintervalos a lo largo del experimento, con valores que se mantuvieron entre 0.1 y 0.3. La Rata R25 mostró valores similares a los del resto del grupo, con la excepción de las primeras sesiones de la Fase 3 y la de reversión en las que las respuestas se concentraron en los Subintervalos 3, 4 y 5.
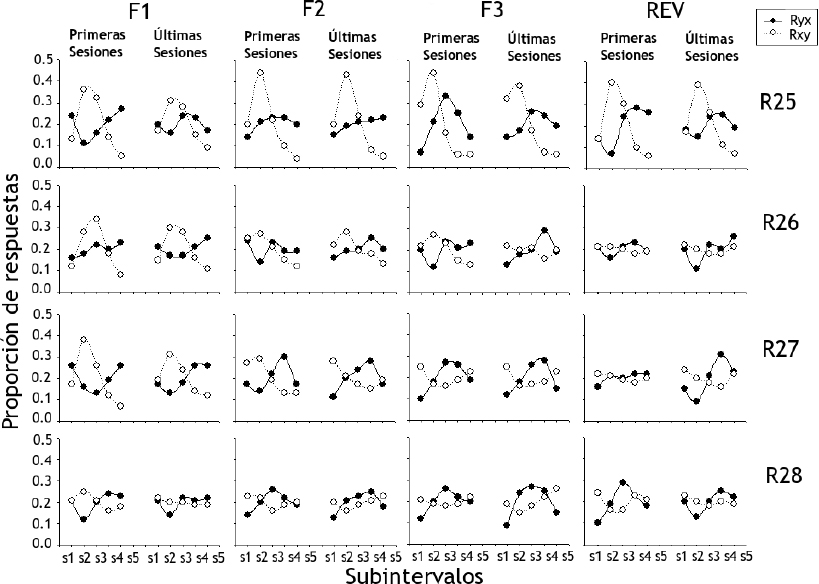
Figura 5. Proporción de respuestas de presión de palanca (Ryx) y de introducción de cabeza al bebedero (Rxy) para las ratas del Grupo 2 según los cinco subintervalos en los que se dividieron los ensayos en las primeras cinco y las últimas cinco sesiones
Donde R es igual a rata y F igual a fase.
En relación con las Rxy, los resultados mostraron que durante la Fase 1 las proporciones con valores más altos fueron las de los Subintervalos 2 y 3, cercanos al 0.4 para todas las ratas, exceptuando la R28. A pesar de que durante las siguientes fases no se encontraron cambios abruptos, conforme transcurrieron las sesiones se observó una tendencia en las proporciones a igualar el valor de 0.2. Este efecto no se encontró en la Rata R25 que mostró una proporción mayor en todas las fases para el Subintervalo 2 con valores cercanos a 0.5.
La Figura 6 muestra la proporción de respuestas de presión de palanca totales (Rs Total) y productoras de agua (Rs Pro) para las ratas del Grupo 1 (ordenada) según la ubicación de las tres palancas en bloques de las primeras y últimas cinco sesiones (abscisa). La proporción de respuestas productoras y totales mostraron valores similares para todas las ratas durante todas las fases. Para la Fase 1, los resultados mostraron una proporción mayor de respuestas productoras y totales para la Palanca 1, alcanzando valores cercanos al 1.0. En la Fase 2 este efecto se replicó para la Rata R21, mientras que en el resto del grupo se observó un decremento con respuestas en las Palancas 2 y 3 de forma más notoria para las Ratas R23 y R24. La Fase 3 se caracterizó por proporciones de valores similares en las tres palancas para las Ratas R23 y R24, con valores próximos a 0.2. Por su parte, las Ratas R21 y R22 mostraron mayor proporción de respuestas en la Palanca 1, pero con valores menores a los mostrados durante las fases anteriores. Al pasar a la fase de reversión se observó el mismo efecto de la Fase 1.
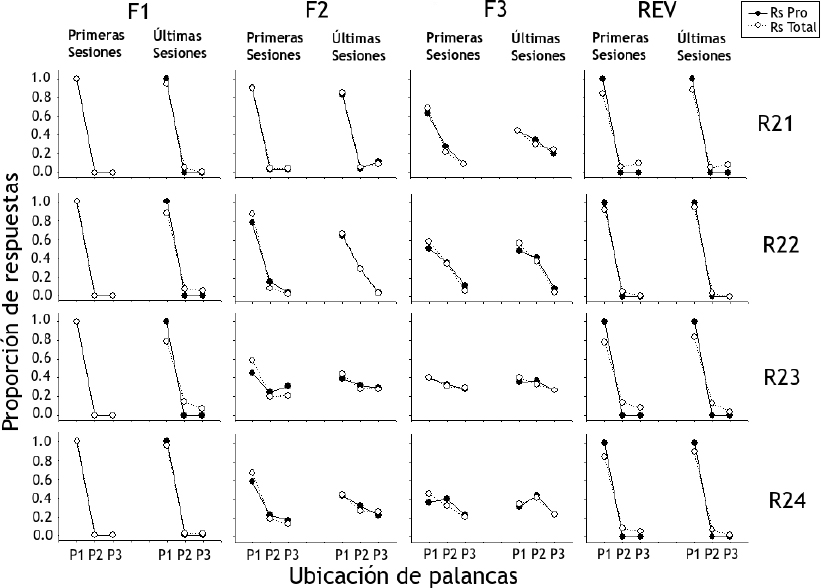
Figura 6. Proporción de respuestas de presión de palanca totales (Rs Total) y productoras de agua (Rs Pro) para las ratas del Grupo 1 según la ubicación de las tres palancas (P1, P2, P3)
Nota. Donde R es igual a rata y F igual a fase.
La Figura 7 muestra la proporción de respuestas de presión de palanca totales (Rs Total) y productoras de agua (Rs Pro) para las ratas del Grupo 2 (ordenada) según la ubicación de las tres palancas en bloques de las primeras y últimas cinco sesiones (abscisa). Al igual que en el Grupo 1, las proporciones de respuestas productoras y totales fueron similares para todas las ratas. Los efectos encontrados en el Grupo 2 concuerdan con el orden en el que las ratas fueron expuestas a las condiciones experimentales de acuerdo con lo observado para el Grupo 1, es decir, exceptuando la Fase 2 los efectos se dieron en orden inverso. Durante las Fases 1 y 2 las proporciones de ambos tipos de respuestas tienen valores similares para las tres palancas, mostrando proporciones ligeramente mayores para la Palanca 1 en comparación con las Palancas 2 y 3. Durante la Fase 3 la proporción de respuestas se concentró en la Palanca 1 para todas las ratas alcanzando valores cercanos al 1.0. La Rata R28 fue la excepción, pues para la proporción de respuestas totales mostró valores menores en comparación con las demás ratas. En la fase de reversión se replicaron los efectos mostrados durante la Fase 1.
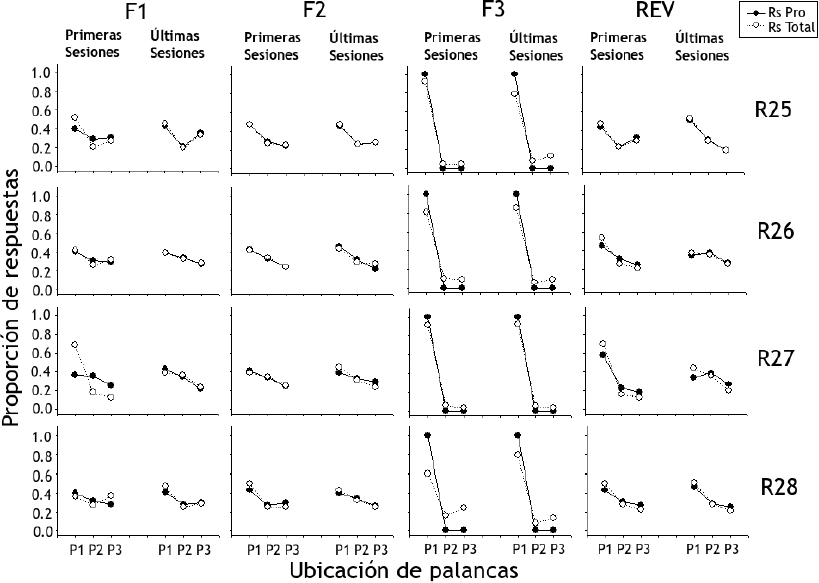
Figura 7. Proporción de respuestas de presión de palanca totales (Rs Total) y productoras de agua (Rs Pro) para las ratas del Grupo 2 según la ubicación de las tres palancas (P1, P2, P3)
Nota: Donde R es igual a rata y F igual a fase.
Discusión
Este estudio se llevó a cabo con el propósito de explorar el efecto de variar el orden espacial de la disponibilidad de producción del estímulo contextualizador (agua) sobre las trayectorias empleadas por el organismo y la ubicación espacio-temporal de sus respuestas (introducción de cabeza al bebedero y de presión de palanca). Con respecto a las trayectorias empleadas, las ratas del Grupo 1 durante la Fase 1 no presentaron de forma clara una proporción mayor por alguna de las trayectorias. Empero, de todas las proporciones la T 12 mostró proporciones mayores, lo que era de esperarse, pues la palanca, la luz y el bebedero se encontraban en la Zona 1. Al pasar a la Fase 2 no se observó una tendencia clara por alguna trayectoria, y en la Fase 3 se observaron efectos similares a los de la Fase 1.
Este hallazgo implica que durante la Fase 3 las ratas se comportaron de forma similar a la Fase 1 en la que la producción del agua fue posible únicamente en la palanca sur (P1) a pesar de que la disponibilidad varió aleatoriamente en las tres palancas. Probablemente, el pasar de una situación en la que las ratas podían anticipar el lugar dónde se produce el agua a otra en la que era imposible hacerlo, promovió la emergencia de un fenómeno parecido al resurgimiento (Epstein, 1983). Empero, en este caso resurge el empleo de una trayectoria previamente extinta.
Para el Grupo 2 los resultados mostraron una tendencia en todas las ratas a emplear en mayor proporción las Trayectorias 12 y 123. Esto sugiere que las ratas, al ser expuestas desde la Fase 1 a una situación en la que era posible producir el agua en las tres palancas, continuaron presentando esta distribución espacial en su actividad, incluso, en la Fase 3 donde la producción de agua sólo fue posible en la palanca sur (P1). Por una parte, este efecto puede ser interpretado como una forma de ajuste diferencial, pues las ratas reprodujeron con su actividad los parámetros espaciales de las ocurrencias con las que los eventos de estímulo fueron programados (Ribes, 2010). Por otra parte, el efecto de arrastre registrado en la Fase 3 puede tomarse como evidencia de desligamiento funcional, pues las ratas permanecieron respondiendo en las tres palancas a pesar de que la disponibilidad se limitó a la palanca sur. En otras palabras, la actividad del organismo se organizó de forma independiente de las propiedades fisicoquímicas concretas de los eventos de estímulo (Ribes; 2015; Ribes y López, 1985).
Respecto de las proporciones de Ryx y Rxy para cada uno de los subintervalos, durante la Fase 1, el Grupo 1 mostró Ryx mayores en los Subintervalos 4 y 5 y Rxy mayores en los Subintervalos 1 y 2, pero este efecto se perdió al cambiar de Fase. En las siguientes fases, las proporciones para ambas respuestas se distribuyeron en todos los subintervalos con valores similares. Posteriormente, en la fase de reversión, se replicaron los efectos de la Fase 1. Por una parte, los efectos encontrados en la Fase 1 y de reversión son similares a lo reportado en estudios que han empleado programas definidos temporalmente (Torres et al., 2015), en lo que respecta a la adecuación o ajuste gradual del comportamiento de las ratas a las contingencias temporales programadas.
Por otra parte, los efectos encontrados en las Fases 2 y 3 parecen indicar que pasar de una situación en la que la producción del agua era espacialmente fija a una en la que la disponibilidad de producción varía en espacio genera una reducción en la diferenciación temporal de los eventos o, lo que desde perspectivas cognoscitivistas, se le ha denominado estimación temporal (Buhusi y Meck, 2009; Matthews y Meck, 2016). Mientras que las ratas en la Fase 1 anticipaban y presentaban las Ryx momentos antes de la ocurrencia del agua y las Rxy durante la entrega del agua, en las Fases 2 y 3 las respuestas se presentaron en todos los subintervalos sin distinción. Los resultados del Grupo 2, apoyan esta proposición ya que desde la Fase 1, en la que la disponibilidad se presentó aleatoriamente en las tres palancas, las proporciones de Rxy y Ryx se distribuyeron de forma similar en todos los subintervalos.
Los resultados obtenidos para las proporciones de respuestas de presión en las tres palancas fueron consistentes entre ratas. Para el Grupo 1 se presentó un efecto de arrastre en las Fases 2 y 3, pues, a pesar de que era posible producir agua en las tres palancas, las Ryx se concentraron en la Palanca 1. Esto no se observó en el Grupo 2; la diferencialidad espacial fue afectada por el orden de las condiciones al que fueron expuestas las ratas.
El efecto de arrastre visto en el Grupo 1 y la mayor acumulación de Ryx en los últimos subintervalos del ensayo en las Fases 1 y de reversión podrían relacionarse con los resultados reportados por Carr et al. (2001) al estudiar el ATL con la tarea de tolva abierta. En ésta, la disponibilidad de producción fue variando en un orden consecutivo a través de distintas palancas y en intervalos estables de tiempo. Con el transcurso de las sesiones los sujetos comenzaron a anticipar el lugar y el momento en que estaría disponible la producción. En una fase posterior, se eliminó el orden de producción, es decir, los sujetos pudieron producir en cualquier palanca, empero, éstos mantuvieron el orden mostrado durante la primera fase (García-Gallardo y Carpio, 2016). Aunque en la tarea de tolva abierta no se emplean señales, como los estímulos visuales del presente estudio, el efecto de arrastre presentado por las ratas implica el mantenimiento de un orden en la distribución espacial de su actividad como efecto de la fase previa.
Como conclusión, a partir de una lógica de complejidad creciente e inclusiva, en la que la función suplementaria incluye relaciones contextuales (González-Becerra et al., 2020; Ribes, 2010, 2018; Ribes y López, 1985), los resultados obtenidos mostraron la estructuración de relaciones contextuales al momento en el que las ratas reprodujeron por medio de su actividad los parámetros espacio-temporales programados para la producción del estimulo contextualizador (agua). En adición, el efecto de arrastre observado en diversas ocasiones puede ser interpretado como evidencia de desligamiento funcional (Ribes, 2015). No obstante, los hallazgos encontrados deben ser tomados con cautela debido a algunas limitaciones en el estudio. Una de éstas consiste en haber empleado modos de representación novedosos, haciendo necesario llevar a cabo otros experimentos similares con el fin de ponerlas a prueba y confirmar la replicación de los resultados. En adición a esto, se debe tomar en consideración algunas características aparatológicas que distinguen a esta investigación, en específico, el haber empleado dispensadores de líquido de tipo gotero, pues esto impide la restricción de acceso al agua por parte de las ratas una vez que la han producido. Finalmente, al comparar los hallazgos aquí obtenidos con los de otros estudios, se debe tomar en cuenta el empleo de ratas hembra como sujetos experimentales, así como el tamaño de las cámaras de condicionamiento ampliadas.
Referencias
Arnau, Jaime (1990). Diseños experimentales en psicología y educación (2a ed., Vol 2). Trillas.
Baum, William & Rachlin, Howard C. (1969). Choice as time allocation. Journal of Experimental Analysis of Behavior, 12, 861-874. https://doi.org/10.1901/jeab.1969.12-861
Buhusi, Catalin & Meck, Warren (2009). Relativity theory and time perception: Single or multiple clocks? Plos One, 4(7), 1-6. https://doi.org/10.1371/journal.pone.0006268
Camacho, Isaac (2017). Teoría de la conducta: una discusión sobre categorías de medida y metodología. Acta Comportamentalia, 25(3), 411-422.
Carr, Jason; Tan, Adrienne; Thorpe, Christina & Wilkie, Donald (2001). Further evidence of joint time-place control of rats' behavior: results from an 'open hopper' test. Behavioural processes, 53(3), 147–153. https://doi.org/10.1016/S0376-6357(01)00138-3
Carr, Jason & Wilkie, Donald (1998). Characterization of the strategy used by rats in an interval time-place learning task. Journal of Experimental Psychology: Animal Behavior Processes, 24, 151-162. https://doi.org/10.1037/0097-7403.24.2.151
Castro, Luis (1977). Diseño experimental sin estadística: usos y restricciones en su aplicación a las ciencias de la conducta (2a ed.). Trillas.
Chamizo, Victoria & Rodrigo, Teresa (2004). Effect of absolute spatial proximity between a landmark and a goal. Learning and Motivation, 35, 102-114. https://doi.org/10.1016/S0023-9690(03)00059-6
Christie, John (1996). Spatial contiguity facilitates Pavlovian conditioning. Psychonomic Bulletin y Review, 3, 357-359. https://doi.org/10.3758/bf03210760
Dembo, Tamara (1930). Zielgerichtetes Verhalten der Ratten in einer freien Situation. Archives Ne´erlandaises de Physiologie de l’Homme et des Animaux, 15, 402–412.
Epstein, Robert (1983). Resurgence of previously reinforced behavior during extinction. Behaviour Analysis Letters, 3, 391-397.
García-Gallardo, Daniel & Carpio, Claudio (2016). Effects of variable sequences of food availability on interval time-place learning by pigeons. Behavioural Processes, 130, 53-64. https://doi.org/10.1016/j.beproc.2016.07.008
González-Becerra, Víctor; Tamayo Tamayo, Jairo; Patrón Espinosa, Felipe; Torres Ceja, Carlos & Ortiz Rueda, Gerardo (2020). Complexity and inclusivity of intrasituational behavior: empirical analysis. Acta Colombiana de Psicología, 23(2), 289-308. https://doi.org/10.14718/ACP.2020.23.2.12
Irigoyen, Juan; Jiménez, Miriam; López, Alfonso & Huerta, José (1999). Análisis de Comportamiento en campo abierto. Revista Sonorense de Psicología, 13(2), 30-46.
Kantor, Jacob (1959). Interbehavioral psychology: A sample of scientific system construction (2nd rev. ed.). Principia Press.
Kupalov, Petr; Voevodina, Olga & Volkova, Valentina (1964). Situational Conditioned Reflexes in Dogs in Normal and Pathological State. Clearinghouse.
Leising, Kenneth; Ruprecht, Chad & Stahlman, David (2014). Modulation of variation by response-reward spatial proximity. International Journal of Comparative Psychology, 27, 326-337.
Maes, Joseph; Fontanari, Laura & Regolin, Lucia (2009). Spatial reorientation in rats (Rattus norvegicus): Use of geometric and featural information as a function of arena size and feature location. Behavioural Brain Research, 201, 285-291. https://doi.org/10.1016/j.bbr.2009.02.026
Matthews, William & Meck, Warren (2016). Temporal cognition: Connecting subjective time to perception, attention, and memory. Psychological Bulletin, 142(8), 865-907. https://doi.org/10.1037/bul0000045
McClearn, Gerald & Harlow, Harry (1954). The effect of spatial contiguity on discrimination learning by rhesus monkey. Journal of Comparative and Physiological Psychology, 47, 391-394. https://doi.org/10.1037/h0059728
Nasser, Helen & Delamater, Andrew (2016). The determining conditions for Pavlovian learning: Psychological and neurobiological considerations. En Robin Murphy y Robert Honey (Eds.). The Wiley handbook on the cognitive neuroscience of learning (pp. 7-46). John Wiley y Sons.
Patrón, Felipe; Torres Ceja, Carlos & Flores Aguirre, Carlos (2014). Efectos de variar la distancia espacial entre los elementos que integran la función suplementaria sobre el ajuste efectivo. Revista Iberoamericana de Psicología: Ciencia y Tecnología, 7(2), 7-17.
Pavlov, Ivan (1928). Lectures on conditioned reflexes: Twenty-five years of objective study of the higher nervous activity (behaviour) of animals. (W. H. Gantt, Trans.). Liverwright Publishing Corporation. https://doi.org/10.1037/11081-000
Pear, Joseph (1985). Spatiotemporal patterns of behavior produced by variableinterval schedules of reinforcement. Journal of the Experimental Analysis of Behavior, 44, 217-231. https://doi.org/10.1901/jeab.1985.44-217
Pear, Joseph & Rector, Brian (1979). Constituents of response rate. Journal of the Experimental Analysis of Behavior, 32, 341-362. https://doi.org/10.1901/jeab.1979.32-341
Ray, Roger & Brown, Douglas (1975). A systems approach to behavior. The Psychological Record, 25, 459-478.
Reebs, Stéphan (1996). Time–place learning in golden shiners (Pisces:Cyprinidae). Behavioural Processes, 36, 253-262. https://doi.org/10.1016/0376-6357(96)88023-5
Rescorla, Robert & Cunningham, Christopher (1979). Spatial contiguity facilitates Pavlovian second-order conditioning. Journal of Experimental Psychology: Animal Behavior Processes, 5, 152-161. https://doi.org/10.1037/0097-7403.5.2.152
Ribes, Emilio (1992). Sobre el tiempo y el espacio psicológicos. Acta Comportamentalia, 0(1), 71-84.
Ribes, Emilio (2007). Estados y límites del campo, medios de contacto y análisis molar del comportamiento: reflexiones teóricas. Acta Comportamentalia, 15, 229-259.
Ribes, Emilio (2010). Teoría de la conducta 2. Avances y extensiones. Trillas.
Ribes, Emilio (2015). El desligamiento funcional y la causalidad Aristotélica: un análisis teórico. Acta Comportamentalia, 23, 5-15.
Ribes, Emilio (2018). El estudio científico de la conducta individual: una introducción a la teoría de la psicología. Manual Moderno.
Ribes, Emilio (2020). El cuerpo de la teoría dela conducta. ¿Teoría de la conducta o teoría de la psicología? En Víctor Alcaraz (Coord.). Festschrift en honor de Emilio Ribes (pp. 33-99). Universidad Veracruzana.
Ribes, Emilio & López, Francisco (1985). Teoría de la conducta: un análisis de campo y paramétrico. Trillas.
Ribes, Emilio; Palacios, Hugo & Hernández, Varsovia (2020). Continuous Measuring of Temporal and Spatial Changes in Rats’ Behavior under Water Temporal Schedules. Psychological Record 70, 267–278. https://doi.org/10.1007/s40732-020-00389-z
Ribes, Emilio & Tamayo, Jairo (2020). Evaluación de diferentes contingencias temporales y espaciales en un ambiente experimental ampliado con cuatro dispensadores de agua. Revista Mexicana de Análisis de la Conducta, 46(2), 84-118. https://doi.org/10.5514/rmac.v46.i2.77875
Rodrigo, Teresa; Gimeno, Elisabet; Ayguasanosa, Meritxell & Chamizo, Victoria (2014). Navigation with two landmarks in rats (Rattus norvegicus): The role of landmark salience. Journal of Comparative Psychology, 128(4), 378-386. https://doi.org/10.1037/a0036544
Saksida, Lisa & Wilkie, Donald (1994). Time-of-day discrimination by pigeons, Columba livia. Animal Learning and Behavior, 22, 143-154. https://doi.org/10.3758/BF03199914
Skuban, William & Richardson, Kirk (1975). The effect of the size of the test environment on behavior under two temporally defined schedules. Journal of the experimental analysis of behavior, 23(2), 271–275. https://doi.org/10.1901/jeab.1975.23-271
Tolman, Edward; Ritchie, Benbow & Kalish, Donald (1946). Studies in spatial learning II. Place versus response learning. Journal of Experimental Psychology, 36, 221-229. https://doi.org/10.1037/h0060262
Torres, Carlos; Monreal, Brenda; Barcénas, Iván & Villamil, Carlos (2015). Efectos de incrementar el intervalo entre entregas de agua bajo diferentes contingencias de disponibilidad limitada. Acta Comportamentalia, 23, 103-115.
FELIPE DE JESÚS PATRÓN ESPINOSA
Doctor en Ciencia del Comportamiento, profesor-investigador en la Facultad de Cien-cias Humanas de la Universidad Autónoma de Baja California. Miembro del Sistema Nacional de Investigadores del Consejo Nacional de Ciencia y Tecnología Nivel 1.
felipe.patron@uabc.edu.mx
https://orcid.org/0000-0003-0340-5751
CARLOS DE JESÚS TORRES CEJA
Doctor en Ciencia del Comportamiento, profesor-investigador en el Centro de Estu-dios e Investigaciones en Comportamiento de la Universidad de Guadalajara. Miem-bro del Sistema Nacional de Investigadores del Consejo Nacional de Ciencia y Tec-nología Nivel 1.
dejesus.torres@academicos.udg.mx
https://orcid.org/0000-0002-4279-8490
CARLOS JAVIER FLORES AGUIRRE
Doctor en Psicología, profesor-investigador en el Centro de Estudios e Investigacio-nes en Comportamiento de la Universidad de Guadalajara. Miembro del Sistema Na-cional de Investigadores del Consejo Nacional de Ciencia y Tecnología Nivel 2.
carlos.faguirre@academicos.udg.mx
https://orcid.org/0000-0002-3085-4029
MAURICIO ORTEGA GONZÁLEZ
Doctor en Psicología, profesor-investigador en la Facultad de Ciencias Humanas de la Universidad Autónoma de Baja California. Miembro del Sistema Nacional de In-vestigadores del Consejo Nacional de Ciencia y Tecnología Nivel 1.
mauricio.ortega40@uabc.edu.mx
https://orcid.org/0000-0002-7349-9186
FORMATO DE CITACIÓN
Patrón, Felipe; Torres, Carlos; Flores, Carlos & Ortega Mauricio (2022). Orden espa-cial de los eventos y distribución de la actividad en la función suplementaria. Quaderns de Psicologia, 24(2), e1731. https://doi.org/10.5565/rev/qpsicologia.1731
HISTORIA EDITORIAL
Recibido: 17-12-2020
Aceptado: 25-06-2022
Publicado: 31-08-2022